- A+
小编属蛇,特别怕蛇,看动物世界,去动物园,有蛇的镜头和画面还想眯着眼看看,怀着这种纠结的心情小编解读了2018年8月1日,国际学术期刊PNAS(《美国科学院院刊》)在线发表中国科学院成都生物研究所李家堂和昆明动物所张亚平等研究团队关于温泉蛇研究的最新成果“Comparative genomic investigation of high-elevation adaptation in ectothermic snakes”
研究背景
青藏高原是地球上海拔最高的高原,平均海拔高度超过4000 m,其相对极端环境的恶劣程度,包括氧化应激,紫外线辐射和极端温度,已导致各种物种的各种适应性反应。对高海拔压力的适应性的生理反应的生物学机制已成为一个多世纪研究的主题。近期在基因组技术方面取得的进展已经为探索适应极端栖息地的遗传基础开辟了新的机会。
温泉蛇属Thermophis属于游蛇科,该属目前包括三个近缘物:青藏高原温泉蛇Thermophis baileyi、四川温泉蛇Thermophis zhaoermii和香格里拉温泉蛇Thermophis shangrila,这三种都是青藏高原的特有物种,生存海拔高度超过3,500米。虽然在温泉附近被发现,但这些物种仍然经历极端的环境条件,包括低浓度的分子氧,高度的紫外线辐射,及每日相对剧烈的温度波动。因此,这些物种对于研究变温动物环境适应性的遗传机制提供了新的思路。
材料与方法
1、研究材料
(1)基因组denovo:青藏高原雌性温泉蛇T. baileyi 1-13:血液样品(基因组denovo);肝脏,大脑,心脏,肺,肌肉和卵巢共计6个组织的混合样品(RNA-seq同源基因预测);
(2)重测序:3只温泉蛇(包含:1只雄性青藏高原温泉蛇Thermophis baileyi、1只雌性四川温泉蛇Thermophis zhaoermii和1只雌性香格里拉温泉蛇Thermophis shangrila)和只2只眼镜蛇(1只雌性大眼睛假眼镜蛇和1只雄性竹假眼镜蛇)(图1);
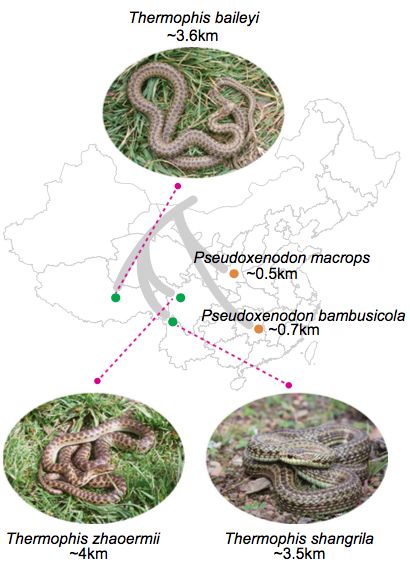
2、文库构建与测序平台
(1)基因组denovo: 双端小片段文库(280 bp和450 bp)和Mate-paired文库 (2 kb, 5 kb, 和10 kb);
(2)全基因组重测序(450 bp);
(3)测序平台:Illumina。
3、分析内容及相关软件
(1)同源聚类(ORTHOMCL v2.0.9);
(2)温泉蛇特有基因统计(BiNGO);GO注释与FDR校正(Benjamini和Hoch- berg);
(3)系统发育树与分化时间计算(ORTHOMCL;MrBayes3.22;PAML4.7);
(4)4DTV分析;
(5)基因家族的收缩与扩张分析(CAFÉ3.1)(6)温泉蛇中共有的氨基酸替代分析(BLASTP;)
(6)FEN1基因功能分析;
(7)EPAS1基因功能析
研究结果
1 、藏温泉蛇基因组denovo
研究中共计获得325 Gb Illumina HiSeq clean reads,测序深度∼185×,预估基因组大小1.76 G(组装了∼1.74 Gb),scaffold N50= 2.41 Mb(比以往组装的蛇的基因组的指标都要高);鉴定了∼791 Mb的重复序列,,其主要由LTR和其它未知的转座因子(TE)组成,占青藏温泉蛇基因组的45.28%,在测序基因组的蛇中其TE转座子的含量是最高的。结合了从头测序和基于同源基因预测策略,共使用来自五个组织的转录组测序数据预测了20,995个蛋白质编码基因。
2、系统进化分析
研究中利用了12个爬行动物基因组和12个非重复脊椎动物基因组的842个单拷贝直系同源基因,进行了贝叶斯系统发育分析(图2),包括基于化石记录的时间校准分析。分子钟分析表明:鳞龙次亚纲(蛇和蜥蜴)和主龙形下纲(海龟,鳄鱼和鸟类)之间的分化发生在二叠纪期间~278.5 Mya,且分化早于二叠纪-三叠纪灭绝事件并与之前的研究报道一致。研究中还讨论了Thermophis属及其近缘属Thamnophis在∼28 Mya发生了分化,Thermophis-Python和Thermophis-Anolis在~71 Mya和~160 Mya发生分化。
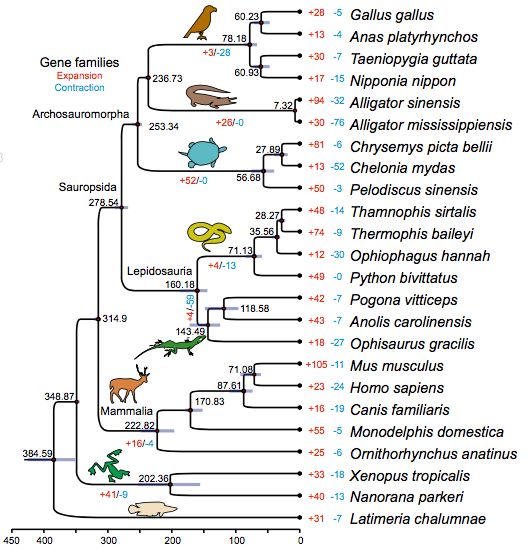
3、基因组家族的进化
利用OrthoMCL聚类,检测到了温泉蛇特异性的92个基因簇(包括354个基因)。这些类别包括感官知觉(嗅觉受体活性,32个基因;G蛋白偶联受体活性,61个基因),对低氧应激的反应(如血红素结合,27个基因;作用于配对供体的氧化还原酶活性,11个基因),以及核糖体相关基因(核糖体的结构成分,29个基因)(图3)。

与其它脊椎动物相比,研究人员还鉴定了青藏温泉蛇中17个显着扩展和两个显着收缩的基因家族(图2)。来自扩张基因家族的基因主要富含L-丙氨酸-2-氧代戊二酸氨基转移酶活性,胆碱酯酶活性,血红素结合,铁离子结合,氧化还原酶活性,单加氧酶活性和碳利用率。两个收缩的基因家族主要由嗅觉受体,基因家族5,亚家族U,成员1和锌指蛋白167组成。
4、正选择基因鉴定与分析
研究中鉴定了青藏温泉蛇中的308个正选择基因(PSGs)。在全基因组上注释了517个基因并涉及“对DNA损伤刺激的响应”这一功能类别。12个基因受到了正向选择,包括:
ERCC6, SMARCAL1, MEIOB, ING4, RBBP5, DMAP1, MSH2, p21, CCNT2, USP7, RBM38和APLF,这些基因大多数位于p53途径的上游或是p53靶标蛋白上(图4)。
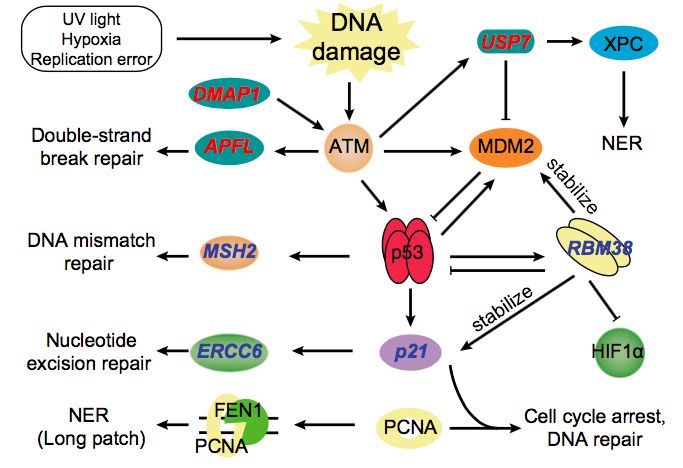
在p53激活途径上游鉴定了3个受选择位点(图4),DMAP1用于ATM(ataxia telangiectasia mutated)的激活,富集到受损伤的部位,与γ-H2AX和复制因子形成复合物,包括增殖细胞核抗原(PCNA)。APLF促进非同源末端连接蛋白复合物的组装和活性,这对维持遗传完整性和修复DNA双链断裂至关重要,及USP7,XPC均参与了DNA的损伤修复。当DNA受到持续的损伤时,刺激p53表达,激活DNA修复蛋白,包括MSH2 , ERCC6, RBM38和p21 (图4)。
青藏温泉蛇在高海拔环境中暴露于紫外线辐射和缺氧状态可能会导致DNA损伤。本研究发现了在紫外线辐射和缺氧状态下受到正向选择的DNA损伤修复基因,暗示了这些基因在低地和高海拔蛇中具有功能性的分化。
5、温泉蛇属共有氨基酸的替换
研究中进而将高海拔温泉蛇中鉴定到了受到正向选择的蛋白序列与9种低海拔蜥蜴和蛇类进行同源蛋白质序列比较。利用SIFT和PolyPhen-2算法来检验温泉蛇与其它物种相比特有的氨基酸替代物的潜在功能效应。在温泉蛇27种蛋白中鉴定了27种特异的氨基酸替换:其中2种蛋白NT5C2和NT5DC3与5'-核苷酸酶活性相关;3种蛋白RNF41, CARNS1和FEN1与活性氧和DNA损伤有功能相关(图5 A);5种蛋白FKB1A, ITSN2, CHST1, WDFY4和IL4RA与免疫相关。缺氧诱导因子(HIFs)(响应细胞环境中可用氧减少的转录因子)超基因家族中,5个基因家族中发现具有11个共有的氨基酸替代物(图5 B)。
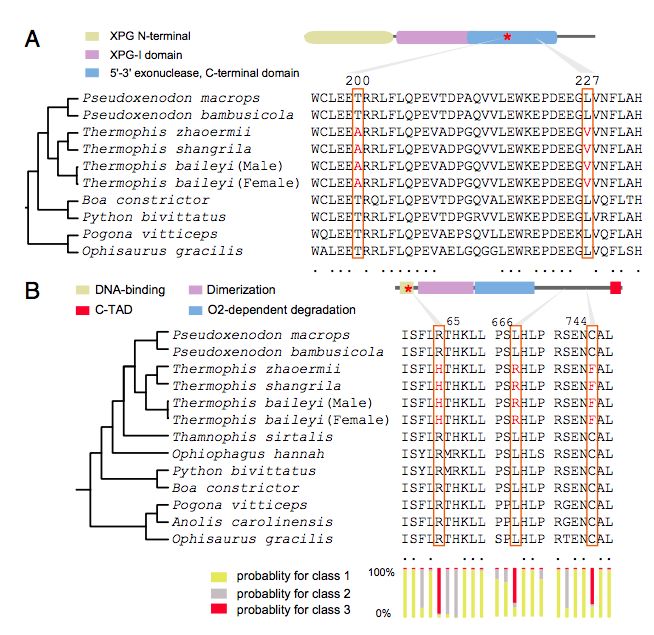
A:蜥蜴和7种蛇FEN1蛋白的变异;
B:3种蜥蜴和9种蛇EPAS1蛋白的变异
6. 紫外线辐射与缺氧的适应性研究
高海拔环境中,紫外辐射增加会导致DNA损伤,DNA损伤反应和修复途径可能显示温泉蛇功能性适应。研究中发现了一些基因与修复途径相关,温泉蛇与所有低地蛇比较,在FEN1中鉴定了两种类氨基酸的替换(图5 A)。FEN1-XPG复合物在体内也显示出显着的NER活性。NER是一种特别重要的切除机制,可以消除紫外线引起的DNA损伤。
在高海拔环境中,温泉蛇属物种不仅暴露于更高的紫外线辐射强度,还暴露于缺氧环境中。EPAS1是HIF超家族的六个成员之一,并已被证明在包括人类和藏獒在内的藏族哺乳动物物种的高海拔地区适应缺氧机制中受到选择,研究中鉴定了温泉蛇属EPAS1中3个共有氨基酸位点的替换,并通过PAML模型揭示了这些蛇中EPAS1基因的显著的正选择作用。
总结
本研究通过比较基因组学分析鉴定了恒温动物与变温动物高海拔适应性的遗传机制。此外组装了第一个高海拔有鳞类温泉蛇属物种青藏高原温泉蛇,为进一步研究变温动物高海拔适应性提供了有利的基因组资源。




